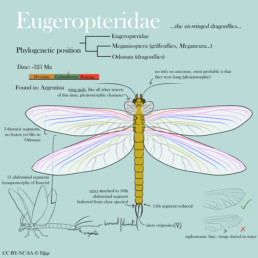
Eugeropteridae are part of the total group Odonatoptera, and in the stem-group to Odonata. They diverged before Meganisoptera (the famous giant « dragonflies » such as Meganeura), and have the characteristic prothoracic wings, a character ancestral to all Pterygota (which are winged insects).
Generalities
As with all insects, the body plan is made of a head (with multiple pieces that I won’t detail), a thorax made of 3 segments, and an abdomen made of 11 segments.
Wings
Wings attach to the three thoracic segments. The first pair are called prothoracic wings, and are in a shape of reduced wing pads (also called « winglets »). Their practical use in vivo is still difficult to assess. Current observations suggest that they were not articulated, and oriented anteriorly. They are strictly homologous to the median and hindwings, but the venation homologies are difficult to identify (and those who claim they can tend to over-interpret). If you’d like to know more, consider to read this very good article from Engel et al. (2013).
On the median- and hindwings, I have represented the main venation, based mainly on the Kirchnerala treintamil specimen (Petrulevicius & Gutierrez, 2016) and with the venation plan from Jacquelin et al. (2018) that applies to all Odonatoptera. Respecting the venation plan is very important, as veins are not placed randomly on an insect’s wings, and their placement is indicative of the evolution of a group.
Quick lesson on insect wings:
It is a common (and old) hypothesis that there is a certain order for wing veins, going from the anterior border to the posterior border, just like fingers on a tetrapod limb, and that they should be homologous among all winged insects. They go as follows:
- Costal vein (C)
- Subcostal vein (Sc)
- Radial vein (R)
- Median vein (M)
- Cubital vein (Cu)
- Anal vein (A)
All veins will have an anterior (A) and posterior (P) branch, thus giving the following pattern : CA, CP, ScA, ScP, RA, RP, etc… Depending on which group we are, we will have a subset of these veins present on the wing, as no insect (yet) has ever been discovered with all of these branches at the same time. Veins and their branches will either disappear or fuse depending on the evolutionary history of the group. This organisation plan is by all means not always consensual. Some authors will add other veins, such as the postcubital vein (PCu) between cubital and anal. It is still a work in progress, and we are waiting for a unified interpretation of wing venation to the scale of all Insects, but until then, wing venation is becoming better and better known, at least at the scale of insect orders.
In this specimen, the costal vein makes up the anterior border of the wing. In red, you have the subcostal veins (anterior and posterior). In pink, you have the radial, anterior (one branch, very long, touches the distal end of the wing). The RP is fused at the base with the MA (purple section), then separates and branches (in pink again). The median (MA and MP) are in blue. In gold, you have the cubital, and in grey the anal.
I have not represented either intercalar nor transverse veins, but they should be added. The placement of transverse is a bit more random though, so there is no need for a guide for that, look at extant insect wings for inspiration.
Wings attach to the thorax with articular sclerites, but there is no consensus that I was able to comprehend, so I abstain from making any suggestion on how to represent them. I have chosen to have 2 big sclerites, loosely inspired from the fossils and extant dragonflies.
Abdominal structures and reproductive organs
All insects have long cerci ancestrally. We attach them to the end of 10th segment, in dorsal position. In extant dragonflies, these cerci are modified to become sturdy claspers with which males graps females by the neck during mating (giving the famous « heart pose » of dragonflies). As cerci are long and thin in Meganisoptera (in Namurotypus to be precise), by simple parsimony they should also be long and thin, preventing any use as a clasper. The terminal filament is a plesiomorphic character, and is present in Meganisoptera (in Namurotypus, again), by parsimony it should thus be present in Eugeropteridae.
About genitalia, if we look at the plesiomorphic state, we can infer quite a few things : the genital orifice is on the anterior and ventral part of the 8th segment ; the ovipositor is attached ventrally, and is short. Ovipositors are made up of multiple parts, and they attach as far back as the 8th segment.
And that’s it!
A personal note
This kind of research is how I view my palaeoart. As a palaeoartist and scientific communicator, it is sometimes hard to be catalogued as simply someone « drawing pretty animals and plants from the past », and that’s it. This is deeply disrespectful to the years of studying, of learning, of doing actual research (in a lab, during both my Master’s degrees, as a qualified volunteer, etc…). I have an academic background, and today this is how I choose to use my knowledge and skills. I refrain from simply tracing photos of fossils, and I talk to/email researchers on precise anatomical features. I hear their pleads about what everybody gets wrong all the time on their favourite taxa, and I try to spread the right message. I don’t mean to undermine the work on palaeoartists who come from a pure artistic background, or those who have never had the chance of such proximity with research. I would just like this work of research to be recognized, even though my technical drawing skills are still far from what I’d like them to reach. One day, I hope my drawing level can draw more attention to the science behind each piece that I produce or supervise during collaborations.
I’d like to thank my secret informator for explaining to me how insects work for years now (and writing excellent blog posts on their own), for giving me bibliography to read and proof-reading and -eyeing this piece. I’d also like to thank Mattia Yuri Messina for reminding me that sources are indeed necessary when we do a serious piece of work, and indirectly motivating me to write this. I probably wouldn’t have done this without that little nudge.
Sources
Engel, M. S., Davis, S. R. & Prokop, J. (2013). Insect Wings: The Evolutionary Development of Nature’s First Flyers. In: Minelli A., Boxshall G., Fusco G. (eds) Arthropod Biology and Evolution. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-36160-9_12
Grimaldi, D. A. & Engel, M. S. (2005). Evolution of the Insects. Cambridge University Press.
Jacquelin, L., Desutter-Grandcolas, L., Chintauan-Marquier, I. et al. New insights on basivenal sclerites using 3D tools and homology of wing veins in Odonatoptera (Insecta). Sci Rep 8, 238 (2018). https://doi.org/10.1038/s41598-017-18615-0
Petrulevicius, J. F., & Gutierrez, P. R. (2016). New basal Odonatoptera (Insecta) from the lower Carboniferous (Serpukhovian) of Argentina. PDF here
Did you like this post? Consider supporting me and my freelance career on my Ko-fi page!
